WB多条带新视角
In my research career of 30+ years, I have encountered hundreds of antibodies that have exhibited multiple banding properties, particularly with tissue lysates. These extra bands bewildered me because it was unclear whether or not these antibodies were specific. It is possible that multiple bands on a Western blot indicate non-specificity, as indicated in Figure 1D in my previous blog post (Blog#1). There are numerous reasons for non-specific binding of an antibody to multiple proteins. One of the frequent challenges is that many proteins, particularly those proteins belong to the same families, can contain the same isotope and result in the development of multiple bands. For example, cardiac α- and β- myosin heavy chain isoforms share 93% sequence identity (Krenz et al., 2003), and the development of an antibody against the shared sequence of the family members will most likely produce multiple bands on a Western blot. Alternatively, multiple bands may result from the detection of a protein during turnover, as protein homeostasis is a dynamic process. All proteins will undergo biogenesis, folding, trafficking, and degradation, however, the rate of turnover varies from protein-to-protein depending on its half-life and cellular processes such as its function and subcellular location (Rahman and Sadygov, 2017). In other words, protein homeostasis (proteostasis) maintains a viable protein pool by constantly undergoing synthesis and degradation (Rahman and Sadygov, 2017). As such, antibodies developed against intact and fragmented portions of a protein will result in multiple bands that often appear as band smearing. Another possibility for multiple bands is a result of post-translational processing such as cleavage or modifications (PTM), including but not limited to, phosphorylation, acetylation, glycosylation, ubiquitination, SUMOylation, succinylation, and methylation. These PTMs are common and indispensable cellular events that are critical for protein maturation and function (Eifler and Vertegaal, 2015; Liu et al., 2016). For example, caspase-1 is activated through self-cleavage and will then cleave pro-IL-1β and pro-IL-18 giving rise to mature interleukin proteins (Malik and Kanneganti, 2017). In cases like these, it is common to detect multiple bands. Lastly, multiple protein isoforms can be derived from a single gene through alternative splicing, a mechanism cells employ to generate a multitude of protein diversity from a limited array of genes (Baralle and Giudice, 2017; Ule and Blencowe, 2019). When these isoforms co-exist within a cell, an antibody may detect multiple bands on a Western blot.
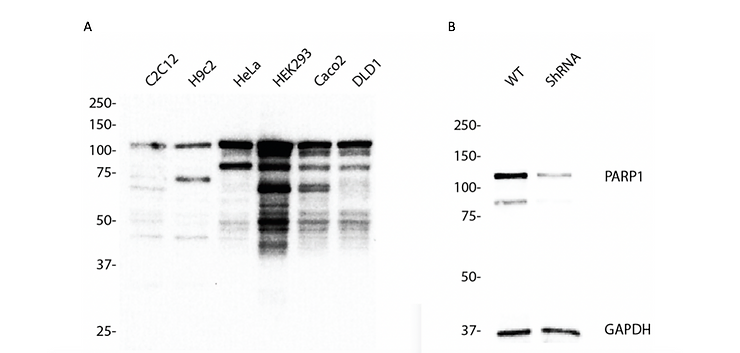 Figure 1. Western blotting analysis using anti-PARP1 antibody (Cat. #6161). PARP1expression in (A) different cell lines and (B) wild type (WT) and PARP1 shRNA knockdown HeLa cells. GAPDH serves as a loading control.
Figure 1. Western blotting analysis using anti-PARP1 antibody (Cat. #6161). PARP1expression in (A) different cell lines and (B) wild type (WT) and PARP1 shRNA knockdown HeLa cells. GAPDH serves as a loading control.
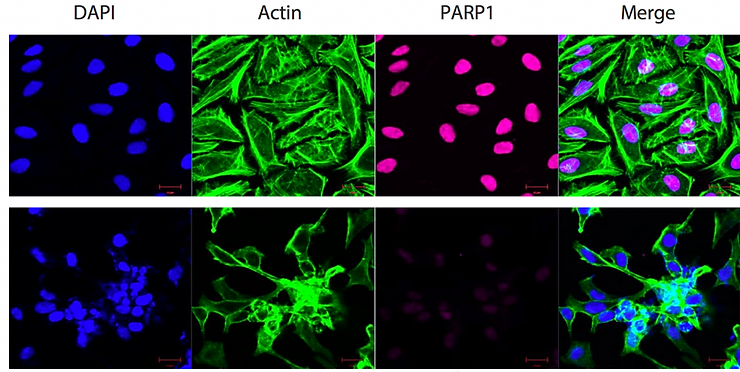
Figure 2. Immunocytochemical staining of HeLa cells with PARP1 antibody. Top panel, wild- type; bottom panel, PARP1 shRNA knockdown. Nuclei were stained blue with DAPI; actin was stained green with Alexa Fluor® 488; PARP1 was stained magenta with Alexa Fluor® 647. Scale bar, 20 μm.
For an example, let’s talk about poly(ADP-Ribose) polymerase 1 (PARP,1 also referred to as PARP) which is a 116 kDa nuclear enzyme involved in DNA damage signaling, chromatin remodeling and epigenetic regulation of gene expression (Ciccarone et al., 2017). The nascent form of PARP1 (116 kDa) can be cleaved by caspase-3 to produce its mature form (89 kDa) (Joshi et al., 2014; Nicholson et al., 1995). From Figure 1A we showed that anti-PARP1 antibody (#6161) recognized both the native and cleaved forms of PARP. The banding pattern indicates that the proteostatic profiles of PARP1 are different across human cell lines and across species (Figure 1A). Additionally, we detected bands below the expected 116 and 89 kDa bands, particularly in HEK293 cells where PARP1 is abundantly expressed (Figure 1A). These extra bands may represent the degraded PARP protein at various proteolytic stages. If our hypothesis is true, knocking down PARP1 should result in the loss of the signal in a Western blot. To this end, we designed a shRNA targeting PARP1 mRNA to lower the level of PARP protein in HeLa cells. Knockdown was confirmed by Western blotting (Figure 1B) as well as immunocytochemistry (Figure 2). The loss of signal in the Western blot and immunocytochemical staining in fixed cells clearly demonstrate that our anti-PARP1 antibody (#6161) is specific.
In summary, multiple bands on a Western blot may indicate that an antibody is specific, or the antibody could be detecting non-specific proteins. Ultimately, a definitive answer is best determined by implementing RNAi and/or knockout strategies, as illustrated by the figures.
References
Baralle, F.E., and Giudice, J. (2017). Alternative splicing as a regulator of development and tissue identity. Nat Rev Mol Cell Biol 18, 437-451.
Ciccarone, F., Zampieri, M., and Caiafa, P. (2017). PARP1 orchestrates epigenetic events setting up chromatin domains. Semin Cell Dev Biol 63, 123-134.
Eifler, K., and Vertegaal, A.C. (2015). Mapping the SUMOylated landscape. FEBS J 282, 3669-3680.
Joshi, A., Mahfooz, S., Maurya, V.K., Kumar, V., Basanna, C.S., Kaur, G., Hanif, K., and Jha, R.K. (2014). PARP1 during embryo implantation and its upregulation by oestradiol in mice. Reproduction 147, 765-780.
Krenz, M., Sanbe, A., Bouyer-Dalloz, F., Gulick, J., Klevitsky, R., Hewett, T.E., Osinska, H.E., Lorenz, J.N., Brosseau, C., Federico, A., et al. (2003). Analysis of myosin heavy chain functionality in the heart. J Biol Chem 278, 17466-17474.
Liu, J., Qian, C., and Cao, X. (2016). Post-Translational Modification Control of Innate Immunity. Immunity 45, 15-30.
Malik, A., and Kanneganti, T.D. (2017). Inflammasome activation and assembly at a glance. J Cell Sci 130, 3955-3963.
Nicholson, D.W., Ali, A., Thornberry, N.A., Vaillancourt, J.P., Ding, C.K., Gallant, M., Gareau, Y., Griffin, P.R., Labelle, M., Lazebnik, Y.A., et al. (1995). Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. Nature 376, 37-43.
Rahman, M., and Sadygov, R.G. (2017). Predicting the protein half-life in tissue from its cellular properties. PLoS One 12, e0180428.
Ule, J., and Blencowe, B.J. (2019). Alternative Splicing Regulatory Networks: Functions, Mechanisms, and Evolution. Mol Cell 76, 329-345.
